Intrón
Definición de intrón
Un intrón es un tramo largo de ADN no codificante que se encuentra entre los exones (o regiones codificantes) de un gen. Los genes que contienen intrones se conocen como genes discontinuos o divididos, ya que las regiones codificantes no son continuas. Los intrones se encuentran solo en organismos eucariotas.

Aquí vemos la estructura de un pre mRNA (o hrRNA) y un mRNA maduro después del procesamiento del mRNA (empalme, la adición de una tapa 5 ‘y una cola poli-A).
Descubrimiento de intrones
Los intrones se descubrieron en 1977 con la introducción de la secuenciación del ADN. Si bien se sabía que las moléculas de ARNm de eucariotas maduras eran más cortas que las transcripciones iniciales, se creía que las transcripciones simplemente se recortaban en los extremos. Cuando se secuenciaron los dos tipos de moléculas, se reveló que este no era el caso; gran parte de la transcripción eliminada provino de regiones internas en lugar de los extremos. Esto provocó una extensa investigación sobre cómo se eliminaron los intrones de las transcripciones y cuál podría ser su función.
Estructura del intrón
En general, los intrones son mucho más largos que los exones; pueden constituir hasta el 90% de un gen y pueden tener más de 10,000 nucleótidos de longitud. Los intrones son frecuentes en los genes; más del 90% de los genes humanos contienen intrones con un promedio de nueve intrones por gen.
Un intrón es un tramo de ADN que comienza y termina con una serie específica de nucleótidos. Estas secuencias actúan como el límite entre intrones y exones y se conocen como sitios de empalme. El reconocimiento del límite entre el ADN codificante y no codificante es crucial para la creación de genes funcionales. En los seres humanos y en la mayoría de los demás vertebrados, los intrones comienzan con 5 ′ GUA y terminan en CAG 3 ′. Hay otras secuencias conservadas que se encuentran en intrones tanto de vertebrados como de invertebrados, incluido un punto de ramificación involucrado en la formación de lariat (bucle).

Aquí vemos una secuencia de consenso para un intrón de vertebrados. El intrón comienza con GUR y termina en un tracto de polipirimidina seguido de YAG.
Función intrón
Si bien los intrones se consideraban inicialmente, y hasta cierto punto todavía se consideran, «ADN basura», se ha demostrado que es probable que los intrones desempeñen un papel importante en la regulación y expresión génica. Como los intrones provocan un aumento en la longitud de los genes, esto aumenta la probabilidad de cruzamiento y recombinación entre cromosomas hermanos. Esto aumenta la variación genética y puede resultar en nuevas variantes de genes a través de duplicaciones, deleciones y mezcla de exones. Los intrones también permiten empalmes alternativos. Esto permite que un solo gen codifique múltiples proteínas ya que los exones se pueden ensamblar de múltiples maneras.
Empalme
Durante la transcripción, la ARN polimerasa copia el gen completo, tanto intrones como exones, en la transcripción de ARNm inicial conocida como pre-ARNm o ARN nuclear heterogéneo (ARNh). Como los intrones no se transcriben, deben eliminarse antes de que pueda producirse la traducción. La escisión de intrones y la conexión de exones en una molécula de ARNm madura ocurre en el núcleo y se conoce como empalme.
Los intrones contienen una serie de secuencias que participan en el empalme, incluidos los sitios de reconocimiento de espliceosomas. Estos sitios permiten que el espliceosoma reconozca el límite entre los intrones y los exones. Los sitios en sí son reconocidos por pequeñas ribonucleoproteínas nucleolares (snRNP). Hay una serie de snRNP involucrados en el empalme de ARNm que combinados crean un espliceosoma.
El empalme se produce en tres pasos:
- Escisión del enlace fosfodiéster entre el exón y la GU en el extremo 5 ‘del intrón. Un snRNP (U1) contiene una secuencia complementaria al sitio de empalme 5 ‘y se une allí para iniciar el empalme.
- Formación de una estructura de lazo o lazo. El extremo libre 5 ‘del intrón se conecta a un sitio de ramificación, una secuencia conservada cerca del extremo 3’ del intrón. Un segundo snRNP (U2) se une al sitio de la ramificación y atrae a U1 para iniciar el lazo. El lazo se forma luego por un enlace fosfodiéster entre el 5 ‘G libre y una A en el sitio de la ramificación.
- Escisión del enlace fosfodiéster entre el segundo exón y el AG 3 ‘del intrón.

Esta figura muestra el empalme de un intrón mediante la formación de un lazo. A continuación, se elimina el intrón dejando los dos exones conectados.
Se desconoce cómo los snRNP y el espliceosoma identifican a qué sitios de reconocimiento unirse dado que los intrones pueden tener miles de pares de bases y hay muchos sitios de empalme crípticos donde las secuencias de reconocimiento se encuentran en otras partes del gen. Se cree que están involucradas ciertas proteínas (por ejemplo, proteínas SR), potenciadores y silenciadores. Los silenciadores de empalme también se han relacionado con enfermedades humanas.
Splicing alternativo
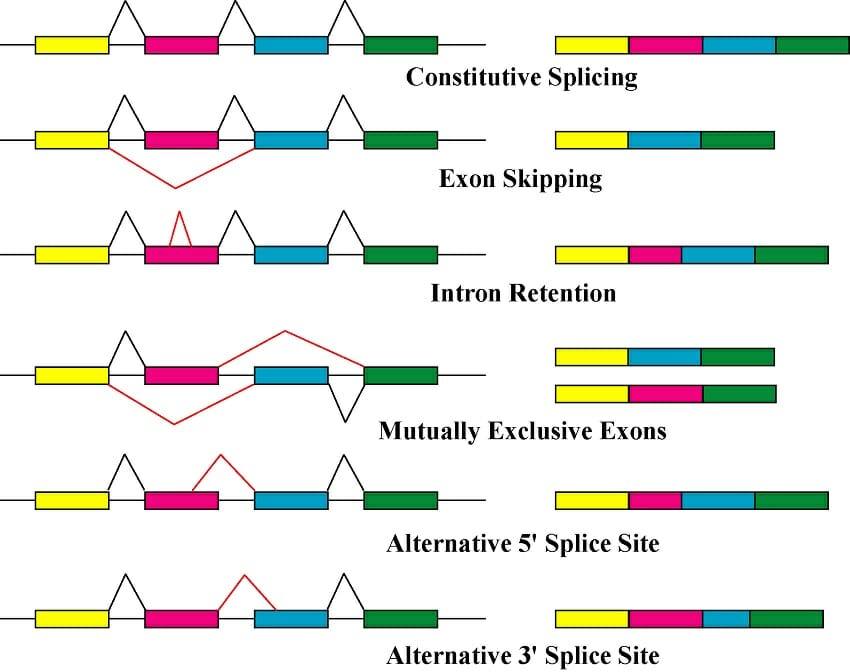
Los intrones y el mecanismo de empalme también permiten productos genéticos alternativos en un proceso conocido como empalme alternativo. Cada gen discontinuo está formado por dos o más exones, lo que permite múltiples formas de ensamblar los exones. El empalme alternativo puede dar como resultado de dos a cientos de ARNm diferentes. El empalme alternativo es común en algunas especies pero raro en otras; se encuentra en más del 80% de los genes humanos, pero solo hay tres casos conocidos en Saccharomyces cerevisiae (levadura).
El empalme alternativo puede ocurrir de varias maneras:
- Salto de exón: uno (o más) exones no se incluyen en el ARNm final
- Retención del intrón: parte del intrón no se empalma correctamente y permanece en el ARNm final.
- Sitio de empalme alternativo: el espliceosoma elimina parte de uno (o más) exones, así como el intrón
ARNr y ARNt
Los intrones también se pueden encontrar tanto en pre-rRNA como en pre-tRNA. Los intrones en los ARNr son raros, y hasta ahora solo se han encontrado ejemplos en eucariotas inferiores. A diferencia de los intrones de otras moléculas, algunos intrones de ARNr tienen una característica única: se auto-empalman. Los intrones auto-empalmados entran en una categoría conocida como intrones del Grupo I. En lugar de depender de una enzima externa para realizar la escisión, los propios intrones actúan como una enzima conocida como ribozima. Las ribozimas se descubrieron en el ciliado Tetrahymena en 1982 y revolucionaron la forma en que los científicos veían las enzimas.
Los intrones en los ARNt son más comunes que los de los ARNr, pero mucho menos prevalentes que en los ARNm, particularmente en los vertebrados (es decir, el 6% de los ARNt humanos).En los ARNt son relativamente cortos, con una longitud de entre 14 y 60 pares de bases. Los intrones forman parte de la estructura de tallo y bucle del ARNt y se unen a una sección del brazo del anticodón. La eliminación de los intrones de pre-ARNt se realiza mediante una única endonucleasa.